SEMINARSKI RAD
Algoritmi koji oponašaju procese u prirodi
Student: Vinko Bradvica
JMBAG: 0036421525
Voditelj: dr.sc.Marin Golub
1. Uvod
U ovom seminarskom radu opisuju se principi i metode rada genetskog algoritma. U prvom dijelu, opisuje se evolucijski proces u prirodi, svojstva jedinki, način pohrane informacija o tim svojstvima, borba za preživljavanje i razmnožavanje. U drugom dijelu opisuju se načini na koje genetski algoritam oponaša metode evolucije iz prirode. Opisuju se različiti načini ocjenjivanja dobrote jedinke, načini selekcije, vrste operatora križanja i mutacije. Nakon toga ukratko se pojašnjava značaj podešavanja parametara genetskog algoritma. U zadnjem dijelu, opisuje se program koji vizualno simulira rad GA u traženju rješenja najboljeg obrasca kretanja jedinke u prostoru.
2. Evolucijski algoritmi
Genetski algoritam je heuristička metoda optimiranja koja oponaša prirodni evolucijski proces, te ga primjenjuje u traženju najoptimalnijeg rješenja zadanog problema. Genetski algoritam oponaša određene značajke evolucijskog procesa, što možemo prikazati u sljedećim koracima - početna populacija jedinki živi, nakon toga prolazi selekciju kako bi se osiguralo da samo najuspješnije jedinke opstanu, zatim one prenose svoj kod na slijedeću generaciju koja se stvara, dok se stara uništava; nova generacija prolazi proces mutacije, a zatim ponavlja te iste korake evolucije kao iprijašnje generacije sve dok se ne ostvari uvjet za zaustavljanje evolucijskog procesa.
Genetski algoritam predložio je dr. John Henry Holland početkom sedamdesetih godina dvadesetog stoljeća. Danas su genetski algoritmi vrlo moćno i praktično oruđe za rješavanje čitavog niza optimizacijskih problema upravo zbog jednostavnosti ideje na kojoj su zasnovani, te jednostavnosti njihove primjene. Unatoč sve većoj primjeni, te povećanom opsegu istraživanja na području genetskih algoritama, rezultati dobiveni na teorijskom području ostaju dvojbeni, a genetski algoritmi su i dan danas u osnovi heurističke metode.
Po načinu djelovanja, genetski algoritmi se ubrajaju u metode usmjerenog pretraživanja prostora rješenja (guided random search techniques) u traženju globalnog optimuma. U istu grupu ubrajamo metode temeljene na sličnim principima kao što su evolucijske strategije i simulirano kaljenje.
Evolucijske strategije koriste realne vektore kao prezentaciju rješenja u kodu, a mutaciju i selekciju kao primarne operatore pretraživanja. Kao što je uobičajeno kod evolucijskih algoritama, operatori se primjenjuju u petlji. Iteracija petlje naziva se generacija. Proces se zaustavlja kad se postigne kriterij zaustavljanja.
Simulirano kaljenje je proces koji kao uzor koristi termodinamičko kretanje materijala prema stanju minimalne energije u postepenom snižavanju temperature kao parametra sustava. Metoda radi s jednim rješenjem od kojega u svakoj iteraciji traži susjedno. Ukoliko se zadovolji uvjet, novo stanje zamjenjuje staro. Čak se može dogoditi i da lošije rješenje zamijeni bolje ako je temperatura dovoljno visoka. Proces kreće sa temperaturom koja ima visoku vjerojatnost prihvaćanja novog rješenja (oko 50%), a zatim se temperatura u svakoj iteraciji smanjuje.
3. Prirodni evolucijski procesi
Evolucija je neprekidan proces u kojemu se jedinke prilagođavaju promjenjivim uvjetima okoliša. Svaka se jedinka može okarakterizirati nekim svojstvima, a ona određuju koliko će ta jedinka biti uspješna u borbi za preživljavanje u trenutnim uvjetima. Najprilagođenija jedinka, s najboljim svojstvima, imat će najviše šansi za preživljavanje i razmnožavanje, dok će slabije jedinke umrijeti ili imati jako male šanse za razmnožavanje. Dakle, jače jedinke će imati više šanse prenijeti dobra svojstva na svoje potomke, dok će loša svojstva odumrijeti zajedno s jedinkama koje nisu dovoljno sposobne.
Svojstva jedinke zapisana su u kromosomima. Jedno svojstvo zapisano je na jednom malom djeliću kromosoma koji se naziva gen. Da bi se pohranila sva svojstva, potrebno je više kromosoma. Kromosomi dolaze u parovima - ukoliko je broj kromosoma potreban da bi se zapisala sva svojstva jednak n, odrasla će jedinka u stanici imati 2n kromosoma, tj. po dva gena za svako svojstvo. Takav par, naziva se alel. U alelu geni mogu biti ravnopravni, kada je svojstvo negdje između svojstva prvog i drugog gena, ili neravnopravni, kada svojstvo određuje dominantni gen.
Jedna dugačka neprekinuta nit deoksiribonukleinske kiseline (DNK) čini devedeset i osam posto kromosoma. DNK se sastoji od dvije spiralne niti građene od fosforne kiseline i šećera, međusobno povezane parovima dušičnih baza, adeninom i timinom odnosno citozinom i gvaninom, kao što je prikazano na slici 3.1. Upravo te dušične baze, nositelji su informacija. Jedna dušična baza može prenijeti četiri informacije, stoga je njezina moć prijenosa informacije ekvivalentna binarnom broju sa dvije znamenke (41 = 22). Ako znamo da čovjek ima oko tri milijarde dušičnih baza u svom genetskom kodu, za zapisivanje tog koda u računalo trebalo bi nam oko 0,75 gigabajta prostora.
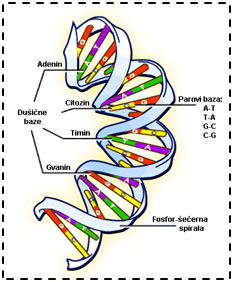
Slika 3.1. - Građa i struktura DNA
Kao što je već spomenuto, broj kromosoma u stanicama odrasle jedinke je 2n. Kako u oplodnji sudjeluju dvije jedinke, spolne stanice imaju haploidan broj kromosoma – n. Rezultantna stanica nakon spajanja spolnih ima 2n kromosoma. Prilikom reprodukcije, dolazi do rekombinacije gena. Ona uzrokuje različitost među pojedincima iste vrste, ali i sličnost s roditeljima. U svakoj novoj generaciji, dobivamo novi skup gena kao kombinaciju roditeljskih - neke će nove jedinke biti lošije, a neke bolje od prethodne generacije. Osim križanja, na promjenu genetske slike neke jedinke mogu utjecati i mutacije – slučajna izmjena genetskog materijala pod utjecajem vanjskih uzroka. One su znatno rjeđe, te je vjerojatnost mutacije od 10-4 do 10-5 ovisno o genu.
4. Genetski algoritam
Genetski algoritam je metoda optimiranja koja imitira prirodni evolucijski proces i primjenjuje ga na apstraktne jedinke. Evolucijski program u svakom svom koraku - generaciji, održava populaciju jedinki. Svaka jedinka je potencijalno rješenje problema koji se obrađuje, predstavljeno podatkovnom strukturom. Te jedinke se nazivaju kromosomi. Kromosom može biti predstavljen brojem, nizom, matricom ili bilo kojom drugom podatkovnom strukturom. Svakoj jedinki se dodjeli određena mjera kvalitete koja se naziva dobrota. Ona se određuje funkcijom dobrote (koristi se i naziv funkcija cilja). Tada se selekcijom odabire skup boljih jedinki iz trenutne populacije, kako bi se iz njih pod utjecajem genetskih operatora reprodukcije stvorila nova populacija. Genetski algoritam prema vrsti selekcije može biti generacijski ili eliminacijski. Genetski operatori reprodukcije dijele se na operatore višeg reda i unarne. Operatori višeg reda kreiraju nove individue kombinirajući osobine različitih jedinki (grupa križanja), dok unarni operatori vrše promjenu nad manjim dijelom genetskog materijala pojedinačne jedinke (mutacijska grupa). Nakon nekog broja izvršenih generacija ili kada se zadovolji uvjet zaustavljanja GA, postupak se zaustavlja, a trenutno najbolje rješenje trebalo bi biti blizu optimuma.
Generacijski genetski algoritam |
4.1. Generiranje početne populacije
Uniformno generiranje početne populacije je postupak kojim se generira početna populacija u kojoj su sve jedinke iste. Kasnije se zahvaljujući mutaciji pojavljuju nova svojstva. Populacija stvorena na ovaj način u početku sporo konvergira rješenju. Neuniformna generacija se može stvoriti na više načina, slučajnim generiranjem jedinki ili stvaranjem populacije od nekih već prije dobivenih dobrih jedinki, koje se onda algoritmom usavršavaju, ili nekom kombinacijom ovih načina.
4.2. Selekcija
Selekcija je proces kojim genetski algoritam čuva dobre, a odbacuje loše jedinke iz populacije rješenja. Selekcijom se odabiru jedinke koje će sudjelovati u reprodukciji, te tako prenijeti svoj genetski materijal na slijedeću generaciju. Kako i loše jedinke mogu sadržavati dobre i korisne gene, potrebno je i njima omogućiti barem minimalnu vjerojatnost za razmnožavanje. Bolje jedinke, naravno, trebaju imati veću vjerojatnost razmnožavanja. Vrste selekcija koje su opisane u ovom radu su jednostavna, linearna i turnirska. Ostale vrste selekcija nabrojane su na slici 4.2. Svaka vrsta selekcije može se primijeniti i u eliminacijskom i u generacijskom GA. Elitizam je postupak koji se uvodi kako bi se najbolja jedinka očuvala te sudjelovala u slijedećoj iteraciji GA neizmijenjena.
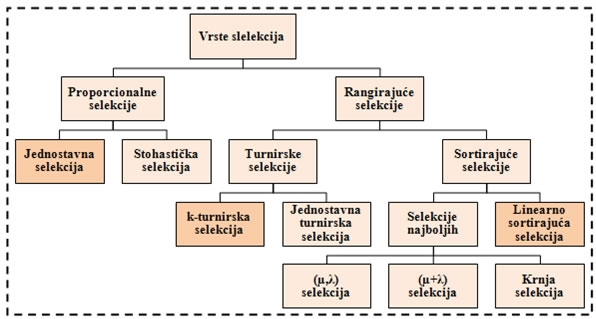
Slika 4.2. - Vrste selekcija, u radu su opisane jednostavna, k-turnirska i linearno sortirajuća
Generacijski genetski algoritam radi s dvije populacije u jednoj iteraciji i opisan je slijedećim pseudokodom:
Generacijski genetski algoritam |
Eliminacijski genetski algoritam za razliku od generacijskog radi samo s jednom populacijom te je opisan slijedećim pseudokodom:
Eliminacijski genetski algoritam |
4.2.1. Jednostavna selekcija
Vjerojatnost izbora jedinke pri proporcionalnoj selekciji proporcionalna je njenoj dobroti. U generacijskom genetskom algoritmu vjerojatnost izbora jedinke računa se tako da se dobrota jedinke podijeli sa ukupnom dobrotom generacije (formula 4.2.). Tako je zapravo vjerojatnost izbora jednaka udjelu njene dobrote u ukupnoj dobroti populacije. Kako je izbor jedinki potpuno slučajan, elitizam se mora uvesti naknadno.
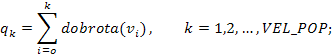
![]()
Postupak jednostavne selekcije opisan je pseudokodom na slici 4.5.
Jednostavna generacijska selekcija |
Eliminacijski genetski algoritam također može koristiti jednostavnu selekciju. U tom slučaju, umjesto funkcije dobrote uvodi se funkcija kazne. (formula 4.3.)
![]()
![]()
Umjesto kumulativne dobrote računa se kumulativna kazna, a jedinka se ne kopira u novu populaciju, već se briše iz trenutne. Ukupnu i kumulativnu kaznu treba računati u svakom koraku, jer jednom izbrisana jedinka više ne pridonosi ukupnoj dobroti. Zbog načina izračuna kazne, elitizam je inherentno ugrađen u selekciju. Algoritam je opisan na slici 4.6.
Eliminacijska selekcija |
Nedostatci jednostavne selekcije su neučinkovitost kod malih razlika u dobroti jedinki, često pojavljivanje duplikata i nemogućnost korištenja negativnih vrijednosti dobrote. Te probleme moguće je ukloniti postupcima translacije ili linearne normalizacije.
Translacija ili pomak je jedan od načina rješavanja nedostataka jednostavne selekcije, kao što su negativne vrijednosti dobrote, velikih vrijednosti i malih razlika dobrote. Rješenje je to da se od vrijednosti dobrote svake jedinke oduzme najmanja vrijednost dobrote iz trenutne populacije. Ukoliko se taj postupak obavi, jedna od jedinki će imati dobrotu jednaku nula i vjerojatnost izbora jednaku nula. Kako želimo da svaka jedinka ima bar malu mogućnost reprodukcije, to se može ispraviti oduzimanjem vrijednosti nešto manje od najmanje vrijednosti dobrote za pozitivne vrijednosti funkcije f(x). Kod eliminacijske selekcije to nije potrebno, jer nam odgovara da najbolja jedinka (najmanja vrijednost funkcije kazne) ima vjerojatnost eliminacije jednaku nula.
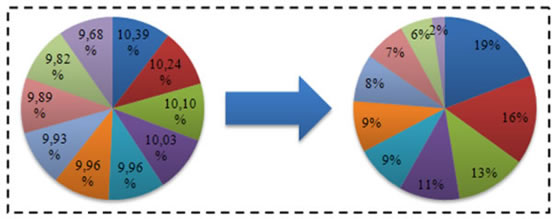
Slika 4.7. – Vjerojatnost izbora jedinki prije i nakon postupka translacije:
dobrota(v(x)) = f(x) – 0.99min(f(x)), prema tablici 4.1.
Linearnom normalizacijom jednostavna selekcija postaje linearno sortirajuća selekcija koja je opisana u slijedećem odjeljku.
4.2.2. Linearno sortirajuća selekcija
U linearno sortirajućoj selekciji vjerojatnost izbora jedinke proporcionalna je poziciji jedinke u poreku jedinki sortiranih prema dobroti. Vjerojatnost selekcije jedinke dana je izrazom 4.5.
![]()
U generacijskom algoritmu, najbolja jedinka ima indeks N, a najgora 1. Za eliminacijsku selekciju koristi se isti izraz, samo najbolja jedinka ima indeks 1, a najgora N. Ova selekcija nema u sebi ugrađen elitizam, te ga je potrebno naknadno implementirati. Ipak, to je vrlo jednostavno napraviti.
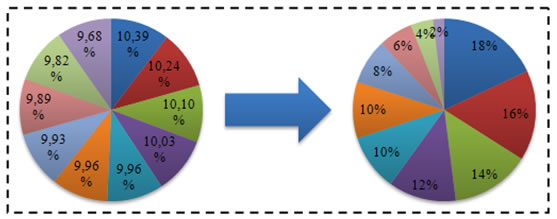
Slika 4.8. – Vjerojatnost izbora jedinki prije i nakon postupka linearne normalizacije,
prema tablici 4.1.
dobrota(v(x)) |
||||||||||
| f(x) | 730 |
720 |
710 |
705 |
700 |
700 |
698 |
695 |
690 |
680 |
| Translacija f(x) – min(f(x)) | 50 |
40 |
30 |
25 |
20 |
20 |
18 |
15 |
10 |
0 |
| Translacija f(x) – 0,99*min(f(x)) | 56 |
46 |
36 |
31 |
26 |
26 |
24 |
21 |
16 |
6 |
| Lin. norm. s korakom 5 i MIN_D =10 | 50 |
145 |
40 |
35 |
30 |
30 |
25 |
20 |
15 |
10 |
| Lin. norm. s korakom 1 i MIN_D =1 | 9 |
8 |
7 |
6 |
5 |
5 |
4 |
3 |
2 |
1 |
4.2.3. K-turnirska selekcija
K-turnirska selekcija je rangirajuća vrsta selekcije koja u svakom koraku za generiranje nove populacije odabire k jedinki iz stare populacije, rangira ih, te najbolju među njima stavlja u bazen za reprodukciju, nad kojim će se onda izvršiti genetski operatori reprodukcije. Eliminacijska turnirska selekcija odabire k jedinki te briše najslabiju od njih iz trenutne populacije, a može ju nadomjestiti djetetom preostale dvije jedinke.
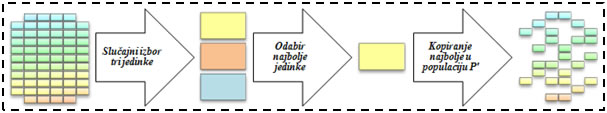
Slika 4.9. – Primjer jednog od koraka generacijske 3-turnirske selekcije.
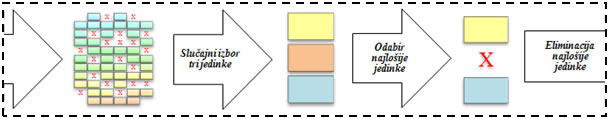
Slika 4.10. – Primjer jednog od koraka eliminacijske 3-turnirske selekcije.
4.3. Genetski operatori reprodukcije
4.3.1. Križanje
Križanje je genetski operator koji imitira prirodni proces križanja (crossover). U njemu sudjeluju dvije jedinke iz populacije, te se križanjem njihovog genetskog materijala dobiva jedna ili dvije nove jedinke. Novonastala jedinka nastaje iz dvije dobre jedinke, koje su prošle selekciju, te bi njihova svojstva trebala biti dobra. Prema tome, nova jedinka bi također trebala biti dobra. Križanje se može izvesti na nekoliko načina. Postupkom križanja se brzo postiže konvergencija optimumu, ali postoji opasnost da on bude lokalni. Najjednostavniji način je križanje s jednom točkom prekida kada se kromosom lomi u jednoj točki te spaja s dijelom drugog slomljenog na istom mjestu.

Slika 4.11. – Primjer križanja jedinki sa jednom točkom prekida.
Križanje sa više točaka prekida obavlja se na sličan način. Jedina razlika je u tome što se kromosomi lome u više točaka, a zatim se dijelovi roditelja A i B naizmjence spajaju. Krajnji slučaj ovog načina križanja je uniformno križanje, tj. križanje s b-1 prekidnih točaka na kromosomu sa b bitova. Kod uniformnog križanja, dijelovi kromosoma se ne spajaju naizmjence već se za svaki gen računa da li je naslijeđen od prve ili druge jedinke koja sudjeluje u križanju, gdje je vjerojatnost nasljeđivanja gena od svakog roditelja jednaka (p=0.5). P-uniformno križanje je uniformno križanje u kojem je vjerojatnost nasljeđivanja pojedinog gena od jednog roditelja drugačija od vjerojatnosti nasljeđivanja od drugog. Vrijednost parametra pi određuje koliko je vjerojatno da će nova jedinka i-ti gen naslijediti od prvrog, odnosno drugog roditelja. Ukoliko vrijedi p = 0, gen se uvijek nasljeđuje od prvog roditelja, a ako je p = 1, onda od drugog. Vrijednost parametra pi = 0.3 govori da će u tri od deset križanja i-ti gen biti od prvog roditelja, dok će u ostalih sedam biti od drugog.

Slika 4.12. – Primjer križanja jedinki sa više točaka prekida.

Slika 4.13. – Primjer uniformnog križanja
4.3.2. Mutacija
Mutacija je unarni genetski operator. Ona djeluje na jedan kromosom, te ga izmjenjuje. Mutacija je operator koji može unijeti novo svojstvo u vrstu ili obnoviti izgubljeno svojstvo. Ukoliko je vjerojatnost mutacije jednaka jedan, tada je algoritam zapravo algoritam traženja slučajnom pretragom prostora rješenja. Što je manja vjerojatnost mutacije, to je manja vjerojatnost nalaska globalnog optimuma. Ukoliko je vjerojatnost mutacije približno nula algoritam će vjerojatno završiti u lokalnom optimumu.
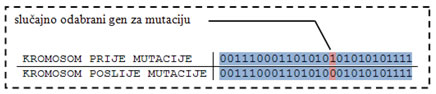
Slika 4.14. – Primjer jednostavne mutacije nad slučajno odabranim genom
Mutacija se može vršiti nad jednim slučajno odabranim genom, nad genima odabranim pomoću slučajno generirane maske, ili nad svim genima – potpuna mutacija. Jednostavna mutacija je najjednostavniji oblik mutacije. Odabranom genu izmjenjuje se vrijednost. Miješajuća mutacija izmiješa pozicije dva ili više odabranih gena. Potpuna miješajuća mutacija slučajno generira vrijednosti odabranih gena, dok invertirajuća miješajuća mutacija invertira vrijednost gena.
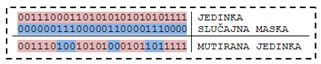
Slika 4.15. – Primjer miješajuće mutacije sa slučajno generiranom maskom

Slika 4.16. – Primjer potpune miješajuće mutacije sa slučajno generiranom maskom
4.4. Parametri algoritma
Parametri generacijskog genetskog algoritma su:
- veličina populacije oznaka VEL_POP
- broj generacija oznaka T
- vjerojatnost mutacije oznaka pm
- vjerojatnost križanja oznaka pc
| parametar | "manje" populacije |
"veće" populacije |
| VEL_POP | 30 |
100 |
| pm | 0,01 |
0,001 |
| pc | 0,9 |
0,6 |
- veličina populacije oznaka VEL_POP
- broj generacija oznaka T
- vjerojatnost mutacije oznaka pm
- broj jedinki za eliminaciju oznaka M
| parametar | "manje" populacije |
"veće" populacije |
| VEL_POP | 30 |
100 |
| pm | 0,01 |
0,001 |
| M | VEL_POP/2 |
VEL_POP/4 |
Za različite vrijednosti parametara, algoritam daje različite rezultate. To se očituje u brzini (može biti brži ili sporiji) te u točnosti rješenja (može dati bolje ili lošije rješenje). Postavljanje djelotvornih parametara zahtjeva izvođenje velikog broja eksperimenata. Parametri mogu biti statički ili dinamički. Statički parametri se definiraju na početku i ne mijenjaju. Dinamički parametri se mogu mijenjati tijekom izvođenja algoritma. Taj proces izmjene parametara može kontrolirati drugi genetski algoritam, može biti funkcija vremena ili broja iteracija odnosno raspršenosti rješenja.
4.5. Simulator genetskog algoritma
U ovom poglavlju nalazi se opis genetskog algoritma - GA Appleta, koji simulira prirodni evolucijski proces. Cilj programa je naći što bolji obrazac ponašanja jedinke - umjetnu inteligenciju.
Svijet jedinki izjelica (eater) podijeljen je u malene kvadratiće. Svaki kvadrat može biti popunjen jedinkom, biljkom ili biti prazan. U jednom se kvadratu ne mogu nalaziti dvije stvari u isto vrijeme. Jedinka, koji ima oblik slova T, "vidi" jedan kvadrat ispred sebe. Ona može vidjeti četiri različite stvari: drugu jedinku, biljku, prazno polje ili zid. Jedinka ima memoriju koja sadrži broj od nula do petnaest. Taj broj predstavlja trenutno stanje jedinke. U svakom intervalu vremena, jedinka može napraviti jednu od ovih akcija:
- Pomaknuti se jedan kvadrat naprijed
- Pomaknuti se jedan kvadrat unatrag
- Okrenuti se na mjestu, 90 stupnjeva ulijevo
- Okrenuti se na mjestu, 90 stupnjeva udesno
Slika 5.1. – GA Applet
U svakom intervalu jedinka također može promijeniti svoje stanje, mijenjajući broj u svojoj memoriji. Ukoliko se pokuša pomaknuti u kvadrat koji je okupiran zidom ili drugom jedinkom, to joj neće biti dozvoljeno. Ipak, ona može u tom intervalu promijeniti stanje. Ukoliko se pomakne u kvadrat koji sadrži biljku, "pojede" ju i osvoji bod. Biljka tada može ili ne mora izrasti negdje drugdje, što ovisi o postavkama podešenim izborniku programa. Na kraju godine (iteracije), dobrota jedinke se određuje prema broju pojedenih biljki plus jedan. GA Applet koristi jednostavni genetski algoritam. Karakteristike su jednostavna selekcija, križanje s jednom točkom prekida i jednostavna mutacija.
Na kraju godine, program ispisuje broj iteracije, srednju vrijednost dobrote populacije i maksimalnu dobrotu. Ukoliko je došlo do nove najveće srednje vrijednosti dobrote, pokraj nje se ispisuje znak '*'. Nakon stotinu godina ili iteracija, rezultati se ispisuju svakih deset godina - desetogodišnji prosjek, maksimalnu dobrotu, te stogodišnji prosjek. Nakon tisuću godina, program ispisuje prosjeke svakih sto godina.
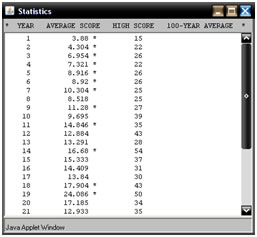
Slika 5.2. – Primjer izlaza GA Appleta
Program može raditi u pet brzina, od kojih najbrža ne prikazuje proces vizualno, što omogućuje brzo napredovanje u evoluciji umjetne inteligencije jedinki.
U programu se mogu podešavati parametri genetskog algoritma:
- VEL_POP–za veličinu populacije može se izabrati bilo koja vrijednost iz skupa A
A = {10, 20, 25, 30, 40, 50, 75, 100}
- pc – za vjerojatnost križanja može se uzeti neka vrijednost iz skupa B
B = {0, 0.1, 0.25, 0.5, 0.75, 1}
- pm – vjerojatnost mutacije može biti neka od vrijednosti iz skupa C
C = {0, 0.0025, 0.005, 0.01, 0.02, 0.03, 0.05, 0.1}
Osim parametara genetskog algoritma, mogu se promijeniti i postavke vezane uz rast biljaka te pozicija na kojoj se jedinke stvaraju (rađaju). Nakon što jedinka pojede biljku, ona može potpuno nestati ili ponovo izrasti – u blizini ili na slučajnoj poziciji. Biljke mogu rasti u nakupinama, nizovima, uz donji rub prozora, uz sve rubove prozora ili na slučajno generiranoj poziciji. Također se može mijenjati i broj biljaka koje izrastu na početku godine (iteracije programa). On se može izabrati iz skupa D.
D = {50, 100, 150, 250, 500}
Eateri se na početku godine mogu stvoriti u gornjem lijevom kutu, sredini prozora, na slučajno generiranoj poziciji ili na poziciji roditelja.
Pokretanjem programa sa različitim postavkama, može se uočiti kako brzina konvergencije prema boljem rješenju ovisi o parametrima genetskog algoritma, dok rješenje prema kojem će algoritam konvergirati ovisi o ostalim postavkama. Program pokrenut s malim i program pokrenut s velikim brojem jedinki u populaciji (VEL_POP) konvergirat će prema sličnom rješenju nakon dovoljnog broja iteracija. Programi pokrenuti s različitim postavkama koje određuju rast biljaka ili pojavljivanje jedinke, neće konvergirati prema istom rješenju.
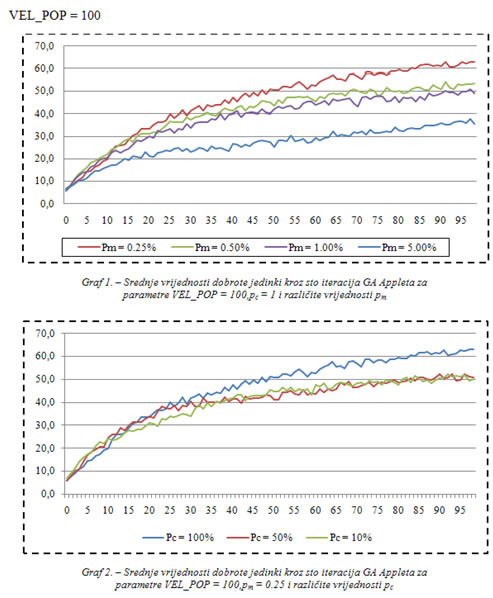
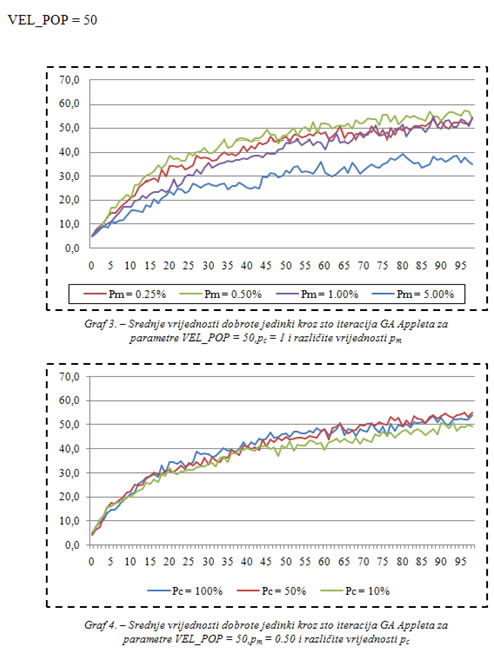
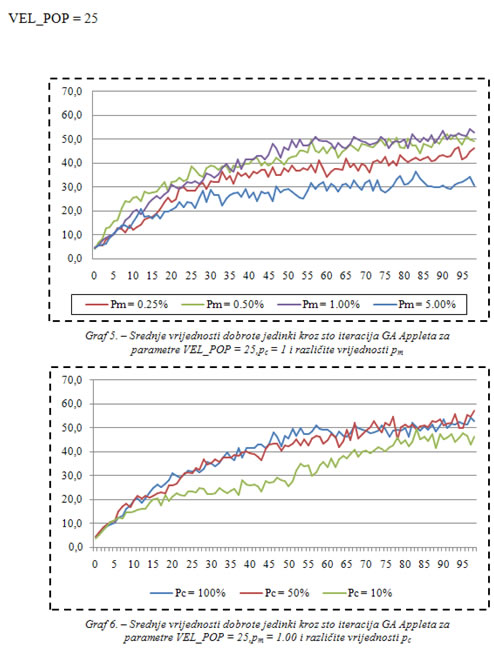
U dodatku se nalaze grafovi koji uspoređuju srednje vrijednosti dobrote populacije za različite vrijednosti vjerojatnosti veličine populacije, mutacije i križanja. Ostale postavke: 500 biljki raste u nizovima, a nakon što ih jedinka pojede pojave se na slučajnoj lokaciji. Jedinke se također pojavljuju na slučajnoj lokaciji.
Najbolje rezultate u prvih stotinu iteracija dale su postavke:
VEL_POP = 100,
pc = 100%,
pm = 0.25%.
Najlošije rezultate u prvih stotinu iteracija dale su postavke:
VEL_POP = 25,
pc = 100%,
pm = 1%.
Iz grafova se može zaključiti da bi postavke VEL_POP = 25, pc = 10%, pm = 5% dale još lošije rezultate u prvih stotinu iteracija, jer se tako ne može postići uniformnost dobrih rješenja zbog male količine prenesenih dobrih, i velike količine novih slučajnih svojstava koja se unose svakom iteracijom posredstvom mutacije.
6. Zaključak
Genetski algoritam je vrlo moćno i praktično oruđe za rješavanje različitih problema optimiziranja. Njegova snaga je u apstraktnosti. Primjenjiv je na velik broj problema, a algoritam nudi puno stupnjeva slobode – nudi poboljšavanje efikasnosti algoritma malim zahvatima. Genetski algoritam daje skup rješenja jednak veličini populacije, a najbolje među njima uvijek bi trebalo biti sasvim blizu optimuma, iako se ne može tvrditi da je sto posto točno. Algoritam se može primijeniti na bilo kojem tipu ili strukturi podataka, što ipak zahtjeva prilagodbu genetskih operatora. Uza svu prilagodljivost genetskog algoritma, i sama ideja algorima je vrlo jednostavna i lako shvatljiva. Ipak, nekad je vrlo teško definirati funkciju dobrote, kako bi dobro opisivala rješenje problema koje tražimo. Stoga se često i problem mora prilagoditi sustavu koji koristimo. Također, podešavanje učinkovitih parametara iziskuje puno vremena i pokusa. Zbog izvođenja velikog broja računskih operacija GA zahtjeva veliku procesorsku snagu i to ga usporava, ali razvojem višejezgrenih i bržih procesora, te ostalih računalnih komponenti, taj problem će biti sve manji.
7. Literatura
-
Golub, Marin, Genetski algoritam: prvi dio. v.2.3. - 27. rujan 2004.,
Zagreb, 2004. -
Golub, Marin, Genetski algoritam: drugi dio. v.2.2. - 4. listopad 2004.,
Zagreb, 2004. -
David, Eck, A Demonstration of the Genetic Algorithm, 24. veljače 2001., http://math.hws.edu/xJava/GA/, 21. travanj 2007.
-
Wikipedia, DNA, http://en.wikipedia.org/wiki/DNA,
21. travanja 2007. -
Wikipedia, GA, http://en.wikipedia.org/wiki/Genetic_algorithm,
21. travanja 2007. -
Wikipedia, Evolution strategies, http://en.wikipedia.org/wiki/Evolution_strategies,
21. travanja 2007.
8. Sažetak
Genetski algoritam je metoda optimizacije koji oponaša prirodni evolucijski proces. GA održava populaciju rješenja, evaluira ih, vrši selekciju nad njima, te ih reproducira i mutira.. Sve informacije o svojstvima neke jedinke u prirodi pohranjene su u jezgrama njenih stanica, u tvorevinama koje nazivamo kromosomima. Kromosomi se sastoje od jedne dugačke niti DNK na kojoj su pohranjene informacije. GA to oponaša, pa svaku jedinku predstavlja upravo jedan kromosom. GA može koristiti bilo koju strukturu i tip podataka u ulozi kromosoma.
Princip rada generacijskog genetskog algoritma temelji se na stvaranju nove populacije od odabranih jedinki iz stare. GA selekcijom odabire uglavnom bolje jedinke, ali i slabije imaju neku šansu preživljavanja. Kada su jedinke kopirane u novu populaciju, nad njima se izvršavaju genetski operatori križanja i mutacije, kako bi nastali njihovi potomci. Time se dobivaju nova rješenja. Eliminacijski genetski algoritam radi samo s jednom populacijom rješenja. Umjesto kopiranja dobrih rješenja, on eliminira loša, te ih nadomješta križanjem i mutacijom preostalih. Potomci mogu biti bolja ili lošija rješenja. GA konvergira k optimumu zahvaljujući križanju, dok se mutacija brine da to ne bude lokalni optumum. Princip selekcije može biti jednostavni, turnirski, eliminacijski ili turnirski eliminacijski. Operatori križanja mogu biti s jednom ili više točaka prekida, dok mutacija može djelovati na jedan ili više bita, tako da im mijenja vrijednost ili zamjenjuje mjesta. Brzina i točnost konvergencije GA uvelike ovisi o parametrima algoritma, kao što su veličina populacije, vjerojatnosti križanja i mutacije, veličine populacije za eliminaciju itd. Optimiziranje tih parametara iziskuje puno vremena i pokusa.
Primjenjivost GA u razvoju jednostavne umjetne inteligencije vizualno dočarava GA Applet. GA Applet je slobodno dostupni simulator rada GA. Jedinkama u tom programu genetski kod određuje uzorak kretanja. Cilj jedinke je pojesti što više hrane. Što je bolji uzorak kretanja, to više hrane će određena jedinka pojesti, te će imati veću vjerojatnost za preživljavanje i reprodukciju.
Genetski algoritam je vrlo jak alat za rješavanje optimizacijskih problema. Prilagodljivost algoritma problemu jedna je od glavnih karakteristika algoritma. Nedostatak algoritma je teškoća definiranje dobre funkcije cilja. Zahvaljujući razvoju jačih računala u budućnosti će se ubrzati izvođenje genetskih algoritama koji traže veliku procesorsku snagu.
DODATAK
grafički prikaz napretka GA u GA Appletu za različite vrijednosti parametara vel_pop, pc i pm